研究内容
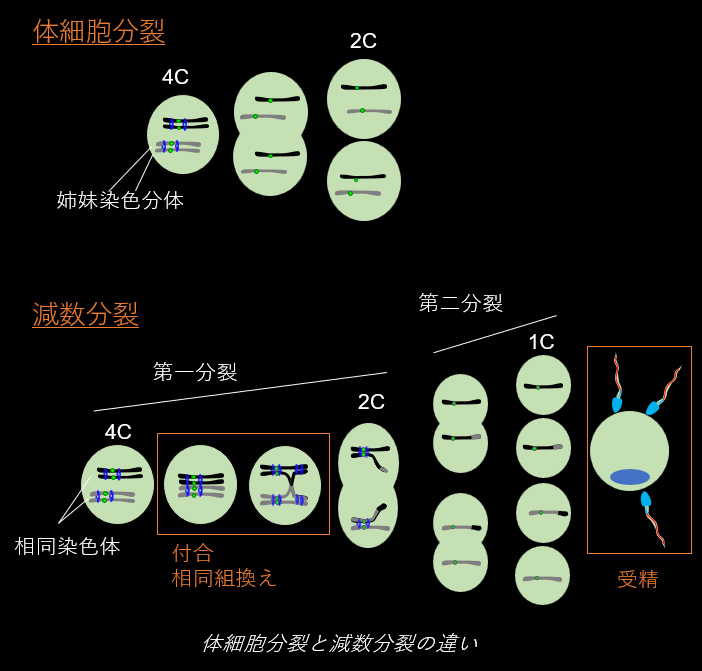
配偶子の特殊性
生き物は個体レベルで寿命を迎えても、生殖細胞において遺伝情報をリフレッシュし、正確に次世代へと引継ぐことで種のレベルでは半永続的に存続可能です。この配偶子の不死性を担保するために、生殖細胞では多くのユニークな生命現象が見られます。まず、減数分裂と呼ばれる特殊な細胞分裂により、染色数が正確に半減されます。その後、受精に特化した細胞形態である精子や卵子への分化が起き、最終的に受精を介して、遺伝情報が次世代へと引き継がれていきます。当チームでは、マウスや線虫の遺伝学、サイトロジー、生化学、等の手法を組合わせ、生殖細胞でのみ見られるユニークな生命現象に焦点をあて、その分子基盤を研究しています。

前期における染色体運動
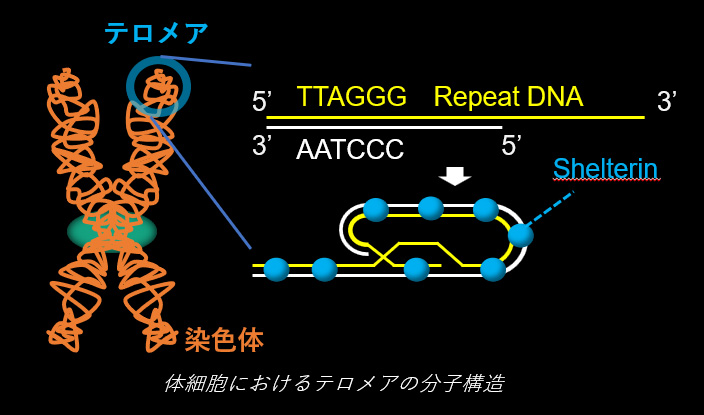
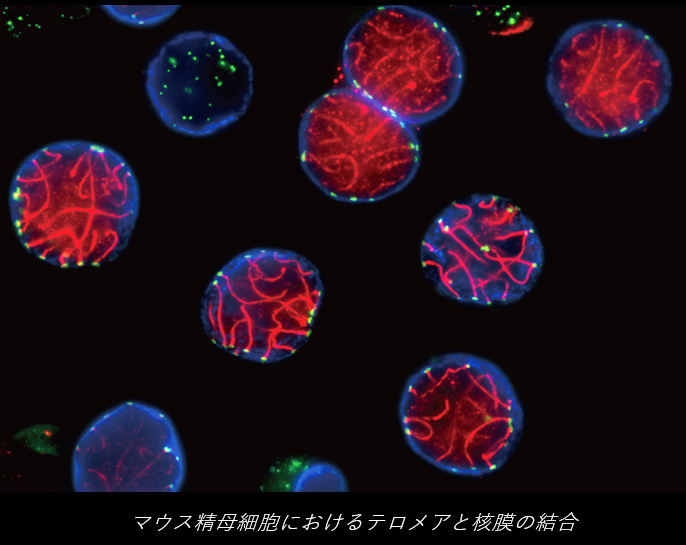
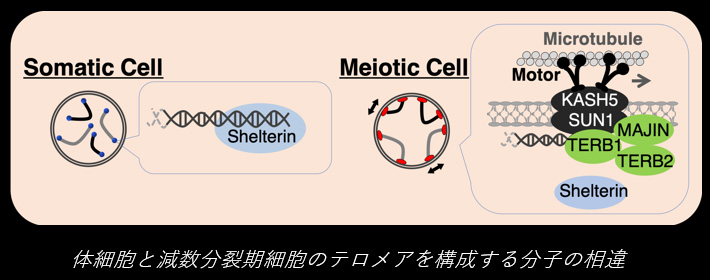
染色体の末端構造であるテロメアは、反復配列からなるDNA鎖と、そこにDNA配列特異的に結合するShelterinタンパク質複合体から成ります。このテロメアの分子構造は普遍的なもので、DNA長の制御、細胞寿命の制御、DNA修復経路の抑制など、生命機能に必須な役割を担っています。近年我々の研究により、減数分裂期前期の精母細胞・卵母細胞では、このテロメアDNAにTERB1-TERB2-MAJINという特殊なタンパク質複合体が結合すること、これによりテロメアDNAが核膜を形成する脂質二重膜へと物理的に連結されることが明らかになりました。
核膜に連結されたテロメアは、核膜を貫通する膜タンパク質複合体であるSUN1-KASH5を呼び込みます。 SUN1-KASH5 は核膜を隔てた細胞質側で、微小管モータータンパク質と結合します。これにより、細胞質モータータンパク質の駆動力が核内の染色体へと伝達され、染色体は核内をダイナミックに動き回ります。この運動は、減数分裂特異的に相同染色体が互いにペアを認識し、対合する過程に必須なものであることが明らかとなりました。
相同組換え
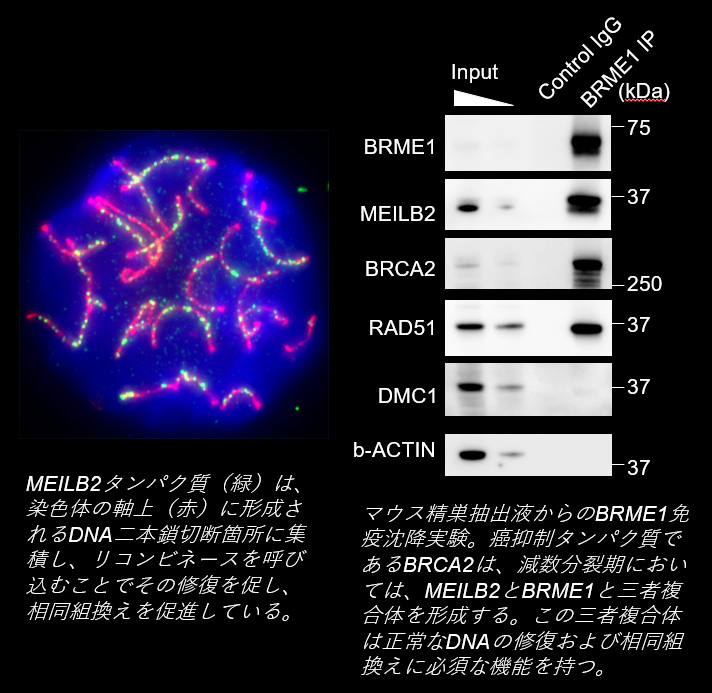
相同染色体のペアがテロメアを介した運動により互いに近接すると、次は相同組換えにより、DNA鎖が交換され、相同染色体はより強固な物理的な結合を獲得します。この物理的な結合はキアズマと呼ばれ、減数第一分裂期に相同染色体が分配されるその時まで、相同染色体のペアを物理的に繋ぎ止める役割を持ちます。我々は、有名な癌抑制タンパク質であるBRCA2が、減数分裂期特異的な相互作用因子であるMEILB2とBRME1と三者複合体を形成することで、リコンビネース(RAD51やDMC1)と呼ばれるDNA二本鎖切断の修復に関わるタンパク質郡をDNA上に呼び込む機能を持つこと、そしてこれにより、正常な相同組換えを制御していることを示しました。
線虫を使い種の存続の分子機構に迫る
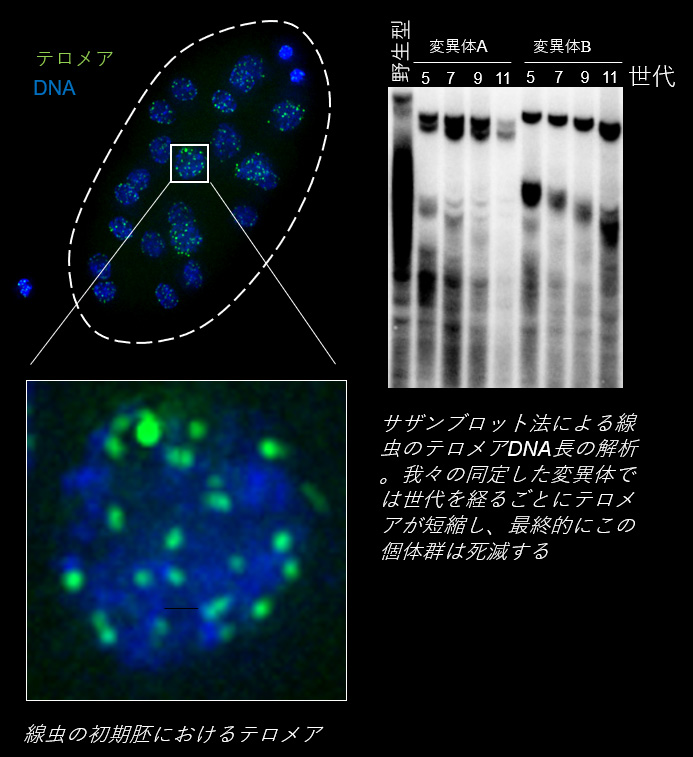
染色体の末端にあるテロメアDNAは細胞分裂のたびに短くなります。そして、限界まで短くなると細胞は分裂を停止します (ヘイフリック限界)。このためテロメアDNAの長さは細胞の寿命を規定すると考えられています。
さて、初めに述べたように、生き物には、個体レベルでは寿命が存在しますが、種としては半永続的に存続可能です。これをテロメアレベルで考えると、生き物には、世代を超えてテロメアDNAを伸長し、種のレベルでテロメア長を未来永劫一定に保つ(恒常性を担保する)メカニズムが備わっていると考えられます。そしてその秘密は次世代にDNA分子を引き渡す細胞である生殖細胞にあるに違いありません。
半永続的に細胞分裂が可能な幹細胞や癌細胞などの特殊な細胞では、テロメレースやALT (Alternative lengthening of telomeres)の活性化によるテロメアDNAの伸長が起こります。生殖細胞でもやはりテロメレースの活性化が起こることが知られていますが、その分子機構はあまりわかっておりません。
世代を超えたテロメア長制御の解明には、世代時間の短いモデル生物を使うのが得策です。例えばポピュラーな哺乳動物のモデル生物であるマウスの場合、その世代時間は約3ヶ月ほどです。仮に、テロメアDNAが世代ごとに短くなり10世代後に異常が出ると期待される変異体を解析する場合、目的のマウスが得られるまでに2.5年かかります。もし仮説が間違っていて、得られたマウスに異常が出なかったら膨大な時間をロスすることになります。そこで、本研究室では世代を超えたテロメア長制御のモデル生物として、世代時間が3日と短い線虫を用いています。線虫は我々と同じ多細胞真核生物であり、遺伝子改変が容易で、雌雄同体であるため自家受精で大量に増えるというメリットもあります。